Sistema ATP-APD
Fuentes de energía
- Células fotosintéticas: fuente de energía la luz. Organismos autótrofos
- Células no fotosintéticas: organismos heterótrofos
Independientemente de la fuente, la energía se obtiene degradando moléculas complejas como hidratos de carbono, grasas, proteínas.
Obtención de la energía
Las moléculas complejas tienen un alto contenido en energía potencial que al degradarlas se desprende. Esto es termodinámicamente favorable. Si las reacciones de degradación no están acopladas a otras la energía se pierde en forma de calor, lo que no es útil (los seres vivos son isotérmicos). Se transporta la energía desde donde se produce hasta donde se necesita acoplando los procesos. Si en una reacción δG < 0 es favorable, pero si δG > 0 se acoplan a procesos muy favorables:
δG = δG1 + δG2 δG1 > 0 δG2 < 0
Si δG < 0 es termodinámicamente favorable. La forma de acoplar reacciones es con un intermediario común. Las reacciones favorables producen el intermediario que luego las desfavorables usan. Para que el metabolismo sea versátil el intermediario ha de ser único, el ATP.
Las reacciones de degradación por oxidación producen ATP a partir de ADP y Pi. El ATP es un nucleósido trifosfato derivado de la adenina. Para sintetizar una molécula el ATP se descompone en ADP y ce energía.
Si la oxidación se produce en condiciones aerobias ocurre en dos etapas:
1.- Oxidación del sustrato nutriente: reacciones catalizadas por enzimas deshidrogenasas que eliminan electrones. Como las proteínas están formadas por aminoácidos que no tiene ninguna cadena lateral adecuada para el transporte electrónico, los enzimas necesitan un cofactor para el transporte. Son dos dinucloeótidos, el NAD+ y el FAD. Una vez recogidos los electrones dan lugar a la forma reducida del cofactor, NADH y FADH2. En esta etapa se sintetiza ATP.
2.- Los cofactores son oxidados por el O2 que recoge los electrones y se reduce a H2O. Los cofactores se reoxidan, NAD+ y FAD y se vuelven a usar. Se sintetiza mucho más ATP que en la etapa anterior.
Como el trabajo es continuo pero el aporte de energía no lo es, el ATP ni se almacena ni se transporta, sino que se produce cuando ser necesita. Cuando sobra energía se aumentan las reservas energéticas.
Catabolismo: reacciones que acompañan a los procesos de degradación y obtención de energía.
Anabolismo: a partir de compuestos sencillos se forman macromoléculas mediante aporte de energía.
A las dos juntas se les llama metabolismo. El ATP producido en el catabolismo se quema en el anabolismo.
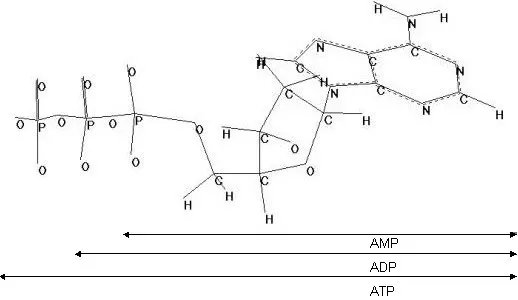
Estructura del ATP
Es un nucleósido trifosfato derivado de la adenina. a pH fisiológico tiene una carga negativa en cada grupo fosfato. Los grupos fosfato se unen mediante enlaces fosfoanhidrilo. Si en lugar de tres tiene dos fosfatos es ADP y si tiene uno es AMP.
Función del ATP
Impulsa reacciones transmitiendo grupos fosfato. Si el aceptor fuera el agua, la reacción de hidrólisis sería:
ATP + H2O ⟶ ADP + Pi
ATP + H2O ⟶ AMP + Pi + pirofosfato
δG°' es δG en condiciones estándar y a pH 7·δG 0 -31 KJ/mol, es muy favorable, el ATP tiene mucha tendencia a ceder Pi o PPi.
δG°' = -2·3·R·T·log Kequilibrio
Al subir Kequilibrio δG se hace muy negativa, por lo que la constante debe ser grande para que cuando participe pueda impulsar otras reacciones.
Kequilibrio = [ADP]·[Pi]/[ATP]
Kequilibrio es tan grande porque influyen dos factores:
- Al perder un Pi tiene menos cargas negativas por lo que es favorable
- El ADP y el Pi están estabilizados por formas resonantes
Requisitos para poder ser moneda energética
La ruptura debe ser favorable y la molécula estable (las transformaciones deben ocurrir a poca velocidad). La energía de activación de ATP + H2O ⟶ ADP + Pi es grande por lo que espontáneamente no ocurre. No hay enzimas que hidrolicen ATP porque la energía se pierde en forma de calor. Si en lugar de agua es otra molécula el sustrato se fosforila.
Hay dos maneras de activar un proceso:
- Que el sustrato forme un derivado con parte de la molécula dando AMP - sustrato
- Que el sustrato se fosforile
Los procesos evolucionan hacia el equilibrio, cuanto más lejos de él más energía dará la transformación.
δG = δGfinal - δGinicio δGfinal = δGequilibrio
δG' depende de las concentración de reactivos y productos en ese momento.
ΔG' = ΔG°' + 2,3·R·T·log ([ADP]·[Pi]/[ATP]).
En condiciones celulares la cantidad de energía que se obtiene al transformar ATP es mucho mayor que δG°'. La relación ATP/ADP está muy lejos de lo que gobierna el equilibrio y la célula gasta mucha energía en mantenerlo así porque δG será mucho más grande.
El ATP es capaz de impulsar una reacción con δG > > 0 porque participa estequiométricamente en ella. Como la reacción es muy favorable es capaz de impulsar la reacción tanto como lejos está del equilibrio.
El sistema ATP - ADP en la célula
Para sintetizar ATP por fosforilación del ADP tenemos acoplando al catabolismo:
ADP + Pi ⟶ ATP + H2O
Sintetizar ATP requiere mucha energía procedente de reacciones de oxidación. Al usar ATP se descompone en ADP ó AMP. Si sólo existieran estas reacciones nos quedaríamos pronto sin ATP, por lo que otras reacciones usan ADP y AMP:
AMP + 2 ATP ⟶ 2 ADP ⟶ 2 ADP + Pi ⟶ ATP
Los enzimas que fosforilan pasan Pi a partir del ATP y se llaman quinasas (adenilato quinasa recupera AMP). Para recuperar PPi:
![]()
GTP, UTP y CTP sirven igual que el ATP:
GTP + H2O ⟶ GDP + Pi ⟶ misma energía que para ATP
Para recuperar el GDP se usa siempre ATP:
GDP + ATP ⟶ GTP + ADP
Fuentes de energía. Estrategias para la síntesis de ATP
Síntesis acoplada a reacciones de degradación por oxidación. En este proceso los cofactores se reducen y se produce energía para sintetizar ATP de ADP + Pi. En la mayor parte de células consta de dos fases:
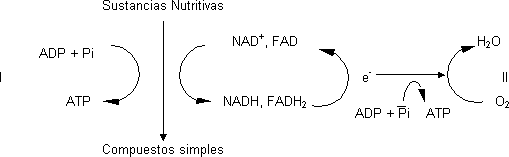
I.- Fosforilación a nivel de sustrato: fosforilación de ADP. Asociada a la fracción soluble de la célula. No requiere O2
II.- Fosforilación oxidativa: se sintetiza mucho más ATP. Síntesis ATP relacionada con la formación de un compuesto fosforilado con potencial de transferencia de grupos fosfato mucho mayor que el ATP para poder cederlo al ADP. Hay mucho compuestos fosforilados pero pocos que puedan cederlo. Estas moléculas se forman como intermediarios de las reacciones metabólicas.
En la reacción de oxidación de aldehído a ácido durante la degradación de la glucosa, con δG grande, el gliceraldehído-3-fosfato por tanto se transforma en 3-fosfoglicerato. Para acoplar la síntesis de ATP se divide la reacción en dos etapas con la formación de un intermediario, el 2,3-BPG:
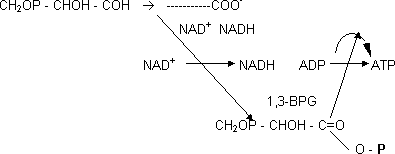
Es inestable, tiene tendencia a ceder el P al ADP.
Hay otros dos compuestos con potencial de transferencia mayor que el ATP:
- Fosfoenolpiruvato (PEP): también interviene en la oxidación de la glucosa
- Fosfocreatina: almacén en el músculo
Membranas transductoras
En la fosforilación oxidativa la cadena está asociada a transporte de electrones. La energía para la síntesis de ATP la proporcionan las reacciones redox durante la cadena de transporte. El proceso está siempre asociado a membranas transductoras de energía que cambian una forma de energía (redox) en otra (ATP). No todas las membranas son transductoras. En las células eucariotas animales las transductoras están en la membrana interna de las mitocondrias, y en las vegetales además en la membrana tilacoidal del cloroplasto. En procariotas la transductora es la membrana plasmática.
Características comunes a todas las membranas transductoras:
Síntesis de ATP acoplada a transporte de electrones. Requisitos:
• Sistema enzimático encargado de la síntesis de ATP (ATP sintasa). Es casi igual para todas las células
• Conjunto de proteínas que formen una cadena de transporte de electrones. Según la célula sea fotosintética o no hay dos tipos:
1) Membrana interna de la mitocondria: encargada de la respiración. Transporte de electrones se realiza a favor de gradiente de potencial (es espontáneo). Los electrones van desde los sustratos (NADH, FADH2) hasta un aceptor final con más tendencia a recibir los electrones. Si hay O2 es el aceptor final
2) Membrana tilacoidal: encargada de la fotosíntesis. Es en contra de gradiente de potencial, no es un proceso termodinámicamente favorable. Sólo funciona con la ayuda de la energía de la luz
Acoplamiento entre la cadena de transporte electrónico y la ATP sintasa.
Por medio de un gradiente de protones. La cadena de transporte y la ATP sintasa están en la misma membrana. Se crea un gradiente a través de membrana por medio de las reacciones redox. La ATP sintasa usa ese gradiente para sintetizar ATP. Además de sintetizar ATP también se usa en muchos procesos.
Autor: Sin datos
Editor: Ricardo Santiago Netto (Administrador de Fisicanet)