Metabolismo de hidratos de carbono
Glicólisis
Transformación de glucosa en piruvato. Participan 10 enzimas y la secuencia de reacciones es la misma para todos los organismos, las diferencias están en la regulación de los enzimas y el destino del piruvato. Cualquier célula la degrada de 2 maneras:
• Fermentación: La glucosa no se degrada completamente porque no se oxida sino que se crean 2 moléculas más pequeñas (de 3 carbonos). Se crea lactato en la fermentación láctica y etanol en la alcohólica. La fermentación no requiere oxígeno. ΔG < 0 = -217 KJ/mol
• Degradación completa hasta CO2. Se necesita la ayuda del O2:
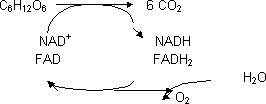
C6H12O6 + 6·O2 ⟶ 6·CO2 ΔG < 0 = -2.810 KJ/mol
No se produce la combustión completa de la glucosa en una sola reacción sino que se hace en muchas etapas liberando energía en porciones utilizables. Hay varias etapas:
Primera etapa o glicolisis:
![]()
Oxidación, intervienen muchos enzimas que recogen todos los electrones del sustrato, para ello necesitan cofactores. Al final de esta etapa tenemos el esqueleto carbonado en forma de CO2 y los electrones recogidos en la deshidrogenasa. No se libera toda la energía, parte la conservan los cofactores.
Segunda etapa:
Los cofactores ceden los electrones al O2 formando H2O. Los cofactores se han de regenerar. La cadena respiratoria transporta los electrones desde los cofactores al O2. La cantidad de energía depende de la tendencia de los cofactores a ceder los electrones y la del O2 para captarlos. Es espontáneo, muy favorable.
Al transportar electrones se bombean protones a través de la membrana:
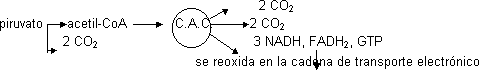
Degradación de la glucosa por proceso llamado glicolisis en el cual la glucosa de 6 C da lugar a 2 piruvatos de 3 C, formando durante esta reacción NADH. El piruvato para degradarse se descarboxila dando 2 de CO2 y 2 de HAc, que se une siempre al acetil-CoA. Se forma NADH. Para degradar el acetil-CoA entra en el ciclo de los ácidos tricarboxílicos (del ácido cítrico o de Krebs).
Cada molécula:
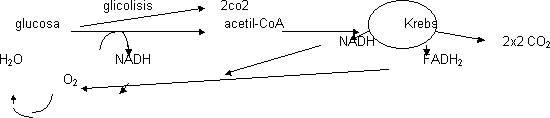
El acetil-CoA se degrada a 2 de CO2. Acoplado al ciclo se crea mucho NADH y algo de FADH2 que ceden los electrones al O2 formando H2O. En las mitocondrias esta cesión se hace en etapas para liberar porciones de energía.
Si el aceptor final es el O2 se trata de respiración aerobia y si es distinto anaerobia. El aceptor final puede ser S que se transforma en SH2 (bacterias del azufre) o el CO2 que pasa a CH4 (bacterias metanogénicas).
Glicólisis
Descripción
Ocurren 10 reacciones a través de las cuales la glucosa se transforma en 2 moléculas de piruvato.
- La glucosa se fosforila nada más entrar en la célula a expensas del ATP y se convierte en glucosa-6-P. Si el P procede del ATP la reacción la cataliza una quinasa que como puede actuar sobre otras hexosas (aunque tiene mayor afinidad por la glucosa) recibe el nombre de hexoquinasa. También se puede fosforilar por otro enzima que no intervenga en la cadena glicolítica. El específico de la glucosa es la glucoquinasa. Tiene una afinidad por la glucosa menor que la quinasa (km más grande). Sólo actúa cuando la concentración de glucosa es muy alta. El paso de glucosa a G6P tiene ΔG' < 0 pero es irreversible
- Para facilitar una segunda fosforilación se pasa a un isómero. La G6P pasa a fructosa-6-fosfato gracias a la fosfoquinasa
- Se fosforila a expensas del ATP gracias a la fosfofructoquinasa convirtiéndose en fructosa-1,6-bifosfato (o FBP). P en C1 y C6. Este paso es irreversible, enzima principal punto de control de la glicolisis. Esta molécula de degrada, no se almacena
- La molécula se parte en 2 de 3 carbonos fosforiladas: gliceraldehído-3-fosfato y fosfato dihidroxi acetona. Enzima encargado aldolasa que lleva a cabo una condensación aldoica. ΔG' > 0, la glicolisis ocurre porque como los productos se consumen se desplaza a la derecha
- Los 2 isómeros pueden transformarse el uno en el otro por medio de una triosa fosfato isomerasa. Este enzima tiene perfección cinética, va todo lo rápido que le llega el sustrato al sitio activo. En el equilibrio el 90% es PDHA. Todo se convierte en Gd3P por lo que la reacción Gd3P ⇔ PDHA está desplazada a la izquierda. El Gd3P recoge todos los carbonos de la glucosa:
![]()
Hasta ahora se han gastado 2 ATP. En la segunda etapa del Gd3P se convierte en piruvato:
- Oxidación del aldehído a ácido 3-fosfoglicerato.-
- La célula guarda la energía de oxidación formando un intermediario acoplado con alto potencial de transferencia de Pi, el 1,3-bifosfoglicerato por medio de la gliceraldehído 3P deshidrogenasa. El enlace ~ tiene gran tendencia a perderse. Ese grupo dirige la síntesis de ATP. Los electrones los recoge el cofactor de la deshidrogenasa, el NAD. Se forma 1 NADH por cada BPG
- Se forma 3-fosfoglicerato por medio de la 3-fosfoglicerato quinasa con gasto de 1 ATP
- El P tiene poco potencial de transferencia por lo que se cambia a la posición 2 por medio de la fosfoglicerato mutasa, siendo el resultado 2-fosfoglicerato
- Se deshidrata por medio de una enolasa dando una forma enólica fosforilada del piruvato, el fosfoenolpiruvato (PEP), que tiene un alto potencial de transferencia del P
- Por medio de la piruvato quinasa (que está regulado) se obtiene piruvato que conserva los carbonos de la glucosa:
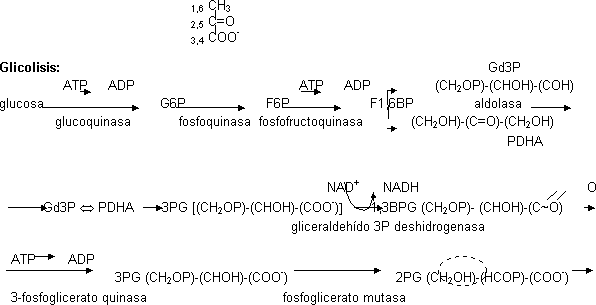
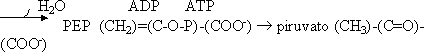
Balance: 1 glucosa = 2 piruvato. Se gastan 2 ATP para los 2 gliceraldehído. En la segunda parte 1 gliceraldehído = 1 piruvato, se producen 2 NADH (1 por cada Gd). Por cada Gd se sintetizan 2 ATP, en total 4 ATP.
![]()
El NADH debe reoxidarse para que la glicolisis no se pare. Si hay O2 es mediante la cadena de transporte electrónico, si no hay otras moléculas se reducen. Depende del entorno del piruvato la manera de oxidarse.
ΔG' = 686 kcal/mol
Si hay O2 suficiente el NADH se reoxida en la cadena de transporte electrónico. Para degradar la glucosa completamente entra en el ciclo de Krebs. Para que el piruvato entre en este ciclo se transforma en acetil-CoA.
Autor: Sin datos
Editor: Ricardo Santiago Netto (Administrador de Fisicanet)