Otros sustratos glicolíticos
Glucógeno
Polisacárido formado por moléculas de glucosa unidas por enlaces glicosídicos. Almacén de glucosa. Para liberar la glucosa se hace una fosforolisis rompiendo con el fósforo y obteniendo el polímero con una unidad menos y una glucosa fosforilada en el C₁. Para incorporarla a la ruta se transforma en G6P mediante mutasa.
Monosacáridos.
Fructosa
Hay dos maneras:
1) Hexoquinasa la fosforila y entra directamente
2) Hexoquinasa tiene más afinidad por la glucosa, si hay mucha no fosforila fructosa. En lugares con mucha glucosa como el hígado se fosforila la fructosa en el C₁ con fructoquinasa obteniendo F1P. F1P ⟶ Gd + PDHA. Sólo queda fosforilar Gd mediante triosa quinasa
Galactosa
Se fosforila mediante quinasa específica formándose galactosa-1-P (epímero de la glucosa). Como debe estar activada se une a UDP dando UDP-glucosa1P, luego se suelta obteniendo glucosa-1-P. En general deben dar intermediarios tras fosforilarse de la glicolisis.
Disacáridos
Sacarosa, lactosa, y por degradación de otros polímeros, maltosa. No entran directamente en la célula sino que se hidrolizan obteniendo los monosacáridos correspondientes:
- Sacarosa: glucosa + fructosa
- Lactosa: glucosa + galactosa
- Maltosa: glucosa + glucosa
Regeneración del NAD⁺
Hay varias posibilidades relacionadas con el destino del piruvato.
- Si hay O₂ funciona la cadena de transporte electrónico y los cofactores se reoxidan
- Si no hay O₂ no hay cadena de transporte electrónico y el piruvato o productos relacionados con él reoxidan el cofactor por fermentación
Fermentación láctica
Transforman el piruvato en ácido láctico.

Puede ir en los 2 sentidos según tengamos NAD⁺ ó NADH.
Balance neto: glucosa + 2 ADP + 2Pi ⟶ 2 lactato + 2 ATP
Se obtiene una pequeña parte de la energía de la glucosa porque no se degrada completamente. El proceso es rentable porque se obtienen 2 ATP sin O₂ y porque se puede recuperar el lactato sintetizando glucosa. Fermentación láctica en el citosol por fosforilación a nivel de sustrato. Los organismos aerobios la utilizan cuando escasea el O₂ y no llega lo suficientemente rápido para reoxidar los cofactores.
ΔG' = -46 kcal/mol
Fermentación alcohólica
Primero el piruvato se descarboxila quedando acetaldehído. Se necesita cofactor porque actúa una carboxilasa, puede ser la TPP (tiamina pirofosfato). Luego se transforma en etanol regenerando NAD⁺
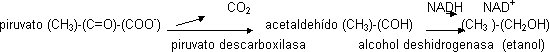
Balance: glucosa + 2 ADP + 2Pi ⟶ 2 CO₂ + 2 etanol + 2 ATP
Transformación del piruvato en acetil-CoA.

Ocurre una reacción de descarboxilación oxidativa, los e⁻ recogidos por el cofactor hacen que pase a NADH (habrá que regenerarlo. La piruvato deshidrogenasa en los eucariotas está en la mitocondria. El piruvato de la glicolisis ocurre en el citosol, si hay O₂ entra en la mitocondria por medio de un transportador específico y el acetil-CoA se libera dentro.
Piruvato deshidrogenasa
Complejo formado por 3 enzimas distintos. Muchas cadenas polipeptídicas (60-80). 3 actividades distintas.
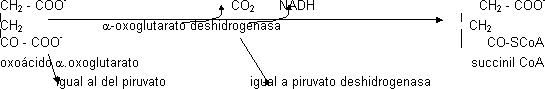
E₁ es la piruvato deshidrogenasa, E₂ es la dihidrolipoil transacetilasa y E₃ es la dihidrolipoil deshidrogenasa. Necesitan 5 cofactores distintos: TPP (E₁), HSCoA, NAD+, FAD (E₃ depende de FAD). El cofactor de E₂, el ácido lipoico, se une covalentemente a la proteína formando un enlace amida por lo que cuando forma parte de la proteína se le llama lipoamida. Además hay 2 enzimas reguladores. La piruvato deshidrogenasa es igual a otro que participa en Krebs pero el sustrato es distinto, el α-oxoglutarato (intermediario en Krebs):
Balance global de la oxidación total de la glucosa:
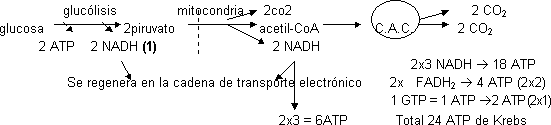
(1) No puede entrar en la mitocondria, manda sólo los e⁻ por medio de una lanzadera.
Total ATP = 24 (Krebs) + 6 (piruvato deshidrogenasa) + 4 (NADH citosol) + 2 (glicolisis a nivel de sustrato) = 36 ATP
Lanzadera del glicerolfosfato
El NADH cede e⁻ al glicerolfosfato que atraviesa la membrana externa. En la interna un enzima lo convierte en PDHA. El cofactor es el FAD que al recoger los e⁻ pasa a FADH₂. Así los e⁻ que estaban en forma de NAD en el exterior pasan al interior en forma de fadh2. Se obtienen 2 ATP porque FADH₂ cede los e⁻ al CoQ (potencial de unión más pequeño). Se produce un gasto de ATP al meter NADH en la mitocondria por lo que al final se obtienen 4 ATP por fosforilación oxidativa.
Otra lanzadera obtiene 3 ATP por cada NADH. Como no implica gasto de energía igual mete NADH que lo saca. Sólo en el corazón.
Gluconeogénesis
Síntesis de glucosa a partir de precursores no hidratos de carbono. Se puede a partir de CO₂ (fotosintéticos) o a partir de restos de 2 (sólo células que tengan el ciclo del glioxilato) ó 3 carbonos. Proceso inverso a la glicolisis. Principales precursores: lactato, glicerol de los lípidos (lípidos son glicerol más ácido graso, no convertibles en hidratos de carbono) y aminoácidos glucogénicos. La glucosa se sintetiza porque se usa como fuente de energía y porque el esqueleto carbonado sirve para construir cosas como ribosas. Hay reservas de glucosa en forma de glucógeno para la energía. Hay células muy selectivas en cuanto al sustrato energético como las cerebrales que sólo admiten glucosa. Si falla el aporte de glucosa se sintetiza para uso de éstas células. La reserva es sólo para 1 día, luego se degradan proteínas.
La gluconeogénesis tiene lugar en el citosol igual que la glicolisis. No son procesos iguales porque en la glicolisis ΔG' << 0, no se puede usar la ruta al revés. Sólo se usan las 7 reacciones que están en equilibrio y no las 3 irreversibles, sino que se da un rodeo.
Glicolisis:
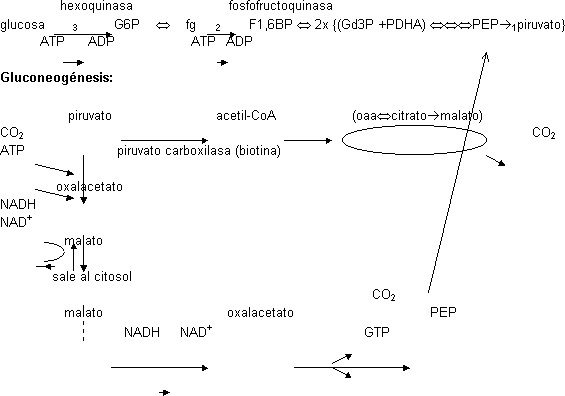
Todos los intermediarios del ciclo de Krebs pueden salir de la mitocondria menos el oxalacetato que no tiene transportador. Se transforma en malato porque esta reacción es reversible en el ciclo del ácido cítrico. El oxalacetato en el citosol se transforma en fosfoenolpiruvato.
Para dar el rodeo se gastan 2 ATP. El CO₂ se gasta y luego se libera y el NAD⁺ se regenera. Si en la otra reacción se producía ATP ahora se gasta, lo mismo para NAD. Para formar una molécula se 6 carbonos se necesitan 2 piruvatos y 2 gliceraldegídos. Se parte siempre de dos piruvatos y el gasto posterior se multiplica por 2. Al llegar a F1,6BP se quita el fosfato mediante una hidrólisis. La fructosa bifosfatasa es el punto de control más importante de la gluconeogénesis:
F1,6BP ⟶ F6
Pi ⟶ H₂O
Al llegar a G6P: [glucosa 6 fosfatasa]
ADP ⟶ ATP
G6P ⟶ glucosa.
H₂O ⟶ Pi
Balance: Si glucolisis y gluconeogénesis ocurrieran simultáneamente se perderían 4 ATP.
Regulación de la glicolisis y gluconeogénesis
Ha de ser conjunta para gluconeogénesis y glicolisis, ha de poner en marcha uno parando el otro. La carga energética es un modulador, cuando está alta activa gluconeogénesis y desactiva glicolisis y viceversa. Son puntos importantes de control la hexoquinasa, la fosfofructoquinasa y la piruvato quinasa.
Regulación de la glicolisis
- - Hexoquinasa: inhibida por G6P que es el producto de la reacción. No fosforilará más si la concentración de G6P es alta. El ATP es un inhibidor también
- - Fosfofructo quinasa: punto más importante de control. Inhibido por ATP y activado por ADP. También inhibido por citrato (primer producto del C.A.C.). Si la concentración de citrato es alta el C.A.C. va más despacio de lo que el sustrato (acetil-CoA) llega para degradar, la concentración de glucosa será más alta. En el C.A.C. se produce mucho NADH y fadh2, para que funcionen se han de reoxidar en la cadena de transporte electrónico creando gradiente de protones. Si el gradiente no se gasta los coenzimas no se reoxidan y el C.A.C. se para
- - Fructosa 2,6 bifosfato: único regulador en glicolisis y gluconeogénesis. Activador para la glicolisis. Se forma a partir de F6P por fosfofructoquinasa₁ que regula también la destrucción

Si la concentración de F26BP es alta es porque hay exceso de glucosa y se activará la degradación.
Regulación de la gluconeogénesis
- Piruvato carboxilasa: primer punto de control. Sólo activa cuando esté presente el acetil-CoA. Si la concentración de ATP es alta activa la gluconeogénesis
- FBPasa: punto más importante de control. Las condiciones celulares determinarán que no funcionen a la misma velocidad porque hidrolizarían ATP. La FBPasa se activa por el ATP y se inhibe por ADP y AMP. Este paso está aún más regulado por estar más controlado

La F26BP regula los dos flujos metabólicos, siendo activa para la PFK e inhibidora para la FBPasa. Cuando aumenta la concentración de FBPasa la glicolisis se activa y se bloquea la gluconeogénesis. Los niveles de F" BP está regulados por los niveles de glucosa. Se forma a partir de F6P por una fosforilación. Estos enzimas varían su actividad por modificación covalente ya que se pueden fosforilar por una quinasa llamada proteín quinasa a, cuando fosforila a la PFK₂ la forma es inactiva y cuando fosforila al otra es activa. La proteín quinasa a se activa cuando los niveles de glucosa son bajos. En ese momento aparece en la sangre una hormona llamada glucagón que al llegar a la célula que puede sintetizar glucosa es reconocida y se activa la proteín quinasa a, de forma que se activará la FBP₂ para que disminuyan los niveles de f26bp, se activa la gluconeogénesis y se inhibe la glicolisis.
- Piruvato deshidrogenasa: está muy regulado porque tiene que ver con la producción de energía. También está regulado por fosforilación (modificación covalente). Hay 2 formas de la enzima, la inactiva es la fosforilada. Si hay mucha energía (concentración de ATP alta) la quinasa estará inactiva. El acetil-CoA es otro modulador positivo, como el NADH. La fosfatasa se ve afectada por la presencia de iones Ca²⁺
Principales sustrato glucogénicos
Lactato: la lactato deshidrogenasa funciona al revés que en la fermentación láctica.
| Lactato | NAD⁺ ⟶ piruvato Lactato | ⟶ NADH ---- ⟶ glucosa. deshidrogenasa |
Glicerol: se obtiene principalmente a partir de las grasas. Es un alcohol de 3 C.
| ATP Glicerol | ⟶ ⟶ Glicerol 3P | ADP Glicerol 3P |
Este glicerol 3P se puede transforma en PDHA que ya es un intermediario de la gluconeogénesis. Estos 2 componentes participan en la lanzadera para introducir NADH en la mitocondria.
Aminoácidos: se incorporan por reacciones de transaminación que dan lugar a los correspondientes oxoácidos que van a la cadena de gluconeogénesis.
Ala ⟶ piruvato, Asp ⟶ oxalacetato, Glu ⟶ α-oxoglutarato.
El requisito para construir glucosa es que sean C₃
Editor: Ricardo Santiago Netto (Administrador de Fisicanet).
Regulación de la glicolisis y gluconeogénesis